En 1975 Daniel Nathans y Hamilton O. Smith descubrieron un tipo de proteínas -las enzimas endonucleasas o enzimas de restricción- que actúan como "tijeras moleculares", cortando la doble cadena de ADN a través del esqueleto de fosfatos sin dañar las bases. El descubrimiento de estas enzimas condujo a dichos microbiólogos al Nobel en 1978 y dio origen a la ingeniería genética.
Las enzimas de restricción son producidas por bacterias como método de defensa contra virus y degradan el ADN extraño.
A su vez, el propio genoma bacteriano está protegido contra sus enzimas de restricción mediante metilaciones (es decir, el agregado de un grupo metilo [-CH3)]) en un átomo específico de ciertos nucleótidos.
Estas moléculas son indispensables para la ingeniería genética, ya que producen fragmentos que se pueden unir entre sí fácilmente (con la ayuda de un "pegamento molecular": la enzima ligasa).
Las enzimas de restricción cortan dejando extremos cohesivos o romos. Los extremos cohesivos son generados cuando la enzima corta las dos hebras asimétricamente, dejando los extremos de cada hebra de simple cadena complementarios entre sí. Por otro lado, los extremos romos son generados cuando la enzima corta las dos hebras por el mismo lugar, generando dos extremos doble cadena.
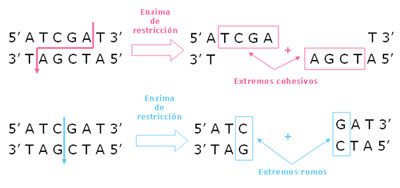
Figura 1. Enzimas de restricción. Este tipo de endonucleasas puede dejar dos tipos de extremos. En el primer caso, el corte genera nucléotidos de simple cadena llamados extremos cohesivos. Estos extremos se pueden unir por medio de otra enzima, la ADN ligasa. En el segundo caso, se generan extremos doble cadena (extremos romos). Estos extremos también pueden ser unidos con la ayuda de una enzima ligasa pero, como no los extremos no son complementarios, la unión será más inespecífica.
Las enzimas de restricción permiten cortar el genoma de cualquier organismo en pequeños fragmentos llamados fragmentos de restricción. La colección de miles o millones de estos fragmentos se llama biblioteca génica.